2011 год № 2
Теоретическая и экспериментальная медицина
Резюме:
Ключевые слова:
Summary:
Key words:
Введение |
 |
 |
Прогестерону принадлежит исключительно важная роль в создании условий для гестационного процесса. В то же время известно, что даже в норме у различных беременных он синтезируется в разных количествах, а при патологическом течении гестации эта вариабельность становится еще большей [2, 5, 6]. Она может усиливаться в результате введения прогестерона женщинам в разные сроки беременности с целью ее сохранения [11]. Вместе с тем, анализ литературы свидетельствует о возможности влияния прогестерона на развитие плода, в том числе его головного мозга. Прогестерону присуще не опасное для плодов обоего пола антиандрогенное действие, проявляющееся через конкурентное подавление им 5α-редукции андрогенов. Это влияние не изменяет прямого действия тестостерона, но регулирует его преобразование в активную форму - дигидротестостерон, которое чрезвычайно важно для половой дифференциации мозга, происходящей у человека в основном во втором триместре беременности [1, 8].
На экспериментальных моделях показано, что мишенями прогестерона являются неокортекс, гиппокамп, гипоталамус и другие отделы мозга. Воздействие на них прогестерона проявляется, в частности, нейрозащитным эффектом [3, 13, 14]. В то же время, сведений о влиянии прогестерона на морфологические показатели развития мозга новорожденных нами не найдено, что и определило цель настоящего исследования - изучить влияние введения прогестерона беременным крысам на развитие головного мозга их новорожденного потомства.
Материалы и методы |
|
|
Исследовались однодневные животные трех групп. Потомство интактных самок составило 1 группу (n=17). Крысята, полученные от самок, которым на 2 и 3 нед. беременности инъецировали персиковое масло (n=11 и n=15 соответственно), составили 2, контрольную группу. В 3 группу было включено потомство самок, которые получали прогестерон на 2 (25 мг/кг) и 3 нед.(25 и 12,5 мг/кг) беременности (n=10, n=25 и n=16 соответственно).
Все животные, потомство которых изучалось, содержались до спаривания и в дальнейшем, после рождения потомства, в условиях одного вивария и получали корм и воду ad libitum. Забивали интактных, контрольных и экспериментальных крысят декапитацией одновременно, в суточном возрасте, определяли массу тела, головного мозга, правого полушария. Левое полушарие крысят фиксировали в жидкости Карнуа. Из переднетеменной (ПТД) и собственно теменной (СТД) долей изготавливали срезы толщиной 7 мкм, которые окрашивали галлоцианином [4]. Гистологическое исследование головного мозга животных сравниваемых групп заключалось в обзорном изучении срезов ПТД, СТД и в изучении архитектоники их неокортекса и ее изменений. На препаратах ПТД и СТД определяли толщину коры и ее слоя I при помощи окуляр-микрометра МОВ-15. Определение плотности расположения нейронов в слоях II, V и гиппокампе проводили при увеличении окуляра х10, объектива х40 в 5 стандартных полях зрения. Статистическую обработку данных проводили с помощью программы Statistica 6.0.
Результаты и обсуждение |
|
|
Введение беременным самкам масла (контроль) нами было осуществлено на 2 и 3 нед. беременности. При этом было установлено, что введение масла на 2 нед. беременности следующим образом отразилось на мозге однодневного потомства. Во-первых, у крысят обоего пола наблюдалось достоверное увеличение массы мозга и полушария. У самцов, кроме того, выявлено статистически значимое увеличение толщины коры СТД, толщины слоя I ПТД и СТД, а также достоверно большая плотность расположения нейронов в слое II ПТД и СТД. У самок достоверные морфометрические отличия включали в себя большую плотность расположения нейронов в слое II СТД и большую толщину слоя I в ПТД и коры СТД (табл. 1).
| Показатели | Интактные | Введение масла беременным самкам | Введение прогестерона беременным самкам (25 мг/кг) | |||
|---|---|---|---|---|---|---|
| самцы (n=10) | самки (n=7) | самцы (n=5) | самки (n=6) | самцы (n=4) | самки (n=6) | |
| Масса тела, г: | 5,4±0,1 | 5,3±0,2 | 5,8±0,2 | 5,6±0,1 | 6,5±0,1*,** | 5,9±0,2* |
| Масса абс., мг: - мозга | 231±7,3 | 234±8,4 | 258±6,7* | 255±3,5* | 282±6,1*,** | 268±7,7* |
| - правого полушария | 72±3,6 | 73±5,9 | 89±2,7* | 87±1,7* | 96±1,3*,** | 88±1,9* |
| Масса отн., мг/г: - мозга | 42,9±1,0 | 44,2±0,9 | 44,8±0,6 | 45,9±0,5 | 43,4±0,9 | 45,8±1,9 |
| - правого полушария | 13,4±0,6 | 13,8±0,8 | 15,4±0,2* | 15,7±0,2* | 14,7±0,1*,** | 15,1±0,4 |
| Толщина, мкм ПТД: слой I | 39,0±2,7 | 43,3±2,0 | 55,3±4,7* | 56,7±2,0* | 56,7±2,9* | 53,6±1,4* |
| - кора | 590±29,6 | 658±8,9 | 637±19,5 | 658±11,8 | 718±11,3*,** | 708±13,4*,** |
| СТД: - слой I | 40,7±3,2 | 51,7±8,4 | 54,8±2,5* | 60,2±2,2 | 61,5±0,3*,** | 59,8±1,8 |
| - кора | 473±3,2 | 491±14,2 | 500±7,3* | 529±7,0* | 537±8,4*,** | 512±13,4 |
| Число нейронов в поле зрения: ПТД: - слой II | 26,1±1,3 | 27,3±1,6 | 29,9±0,9* | 27,9±1,1 | 26,7±0,9** | 26,6±1,0 |
| - слой V | 9,5±0,9 | 9,0±0,5 | 10,1±0,5 | 9,9±0,4 | 8,2±0,2** | 8,5±0,3** |
| СТД: - слой II | 25,8±0,9 | 26,4±1,7 | 29,3±0,4* | 30,1±0,5* | 27,7±0,9 | 28,2±0,9 |
| - слой V | 10,0±0,5 | 10,3±0,5 | 11,1±0,3 | 10,4±0,4 | 9,3±0,4** | 9,9±0,3 |
| - гиппокамп | 24,8±0,8 | 24,3±0,5 | 23,9±0,5 | 25,0±1,2 | 23,9±0,5 | 25,2±0,7 |
Примечания. * - различия с интактными животными статистически достоверны (р<0,05); ** - различия с контролем (введение масла) статистически достоверны (р<0,05).
Обзорное изучение гистологических препаратов мозга крысят сравниваемых групп выявило, что введение масла приводило к увеличению степени стратификации неокортекса (рисунок). Введение масла самкам на 3 нед. беременности также обусловило отличия мозга их однодневного потомства от мозга интактных крысят. У животных обоего пола было найдено достоверное увеличение массы мозга, его полушария и толщины коры СТД. У самок, кроме того, выявлено статистически значимое возрастание плотности расположения нейронов в слое V ПТД (табл. 2).
| Показатели | Интактные | Введение масла беременным самкам | Введение прогестерона беременным самкам | |||||
|---|---|---|---|---|---|---|---|---|
| самцы (n=10) | самки (n=7) | самцы (n=6) | самки (n=9) | 12,5 мг/кг | 25 мг/кг | |||
| самцы (n=6) | самки (n=10) | самцы (n=13) | самки (n=12) | |||||
| Масса тела, г: | 5,4±0,1 | 5,3±0,2 | 5,9±0,1* | 5,8±0,1* | 5,9±0,1* | 5,5±0,1 | 5,6±0,1 | 5,4±0,2 |
| Масса абс., мг: - мозга | 231±7,3 | 234±8,4 | 270±4,8* | 271±2,8* | 269±2,2* | 264,5±3,2* | 248±5,9** | 243±7,3** |
| - правого полушария | 72±3,6 | 73±5,9 | 94±1,8* | 95±1,2* | 90,5±1,6* | 87±2,6 | 77,8±2,3** | 75±2,8** |
| Масса отн., мг/г: - мозга | 42,9±1,0 | 44,2±0,9 | 45,8±1,5 | 46,9±1,1 | 45,5±0,6* | 47,9±0,7* | 44,0±0,5 | 44,9±0,6 |
| - правого полушария | 13,4±0,6 | 13,8±0,8 | 15,9±0,4* | 16,4±0,4* | 15,3±0,4 * | 15,8±0,4* | 13,8±0,3** | 13,8±0,3** |
| Толщина, мкм ПТД: - слой I | 39,0±2,7 | 43,3±2,0 | 52,0±1,0 | 51,8±3,6 | 55,5±1,5* | 55,4±2,9* | 50,5±1,5* | 56,0±3,0* |
| - кора | 590±29,6 | 658±8,9 | 664±13,2 | 662±21,9 | 660±18,0 | 666±9,4 | 658±30,1* | 672±18,0 |
| СТД: - слой I | 40,7±3,2 | 51,7±8,4 | 57,8±1,3 | 55,3±1,9 | 53,0±1,2* | 55,4±1,4 | 56,0±3,0* | 60,5±2,5 |
| - кора | 473±3,2 | 491±14,2 | 556±16,6* | 550±11,6* | 513±23,5 | 538±13,4* | 537±14,5* | 534±21,5 |
| Число нейронов в поле зрения: ПТД: - слой II | 26,1±1,3 | 27,3±1,6 | 24,0±0,2 | 26,5±0,6 | 27,0±0,5** | 26,1 ±0,4 | 25,1±0,1 | 25,2±1,8 |
| - слой V | 9,5±0,9 | 9,0±0,5 | 8,4±0,6 | 11,2±0,4* | 10,8±0,6 | 10,8±0,5* | 8,7±0,7 | 8,4±0,2** |
| СТД: - слой II | 25,8±0,9 | 26,4±1,7 | 26,1±0,5 | 27,6±0,5 | 26,6±1,0 | 27,1±1,0 | 23,3±1,3 | 24,4±0,6** |
| - слой V | 10,0±0,5 | 10,3±0,5 | 10,2±0,2 | 10,7±0,4 | 10,6±0,6 | 10,3±0,5 | 10,2±0,4 | 9,8±0,4 |
| - гиппокамп | 24,8±0,8 | 24,3±0,5 | 23,7±0,5 | 23,6±0,4 | 23,8±0,4 | 25,4±0,3 | 23,2±1,1 | 24,7±0,3** |
Примечания. * - различия с интактными животными статистически достоверны (р<0,05); ** - различия с контролем (введение масла) статистически достоверны (р<0,05).
При обзорном изучении препаратов мозга выявлено возрастание степени стратификации неокортекса по сравнению с интактными крысятами (рисунок).
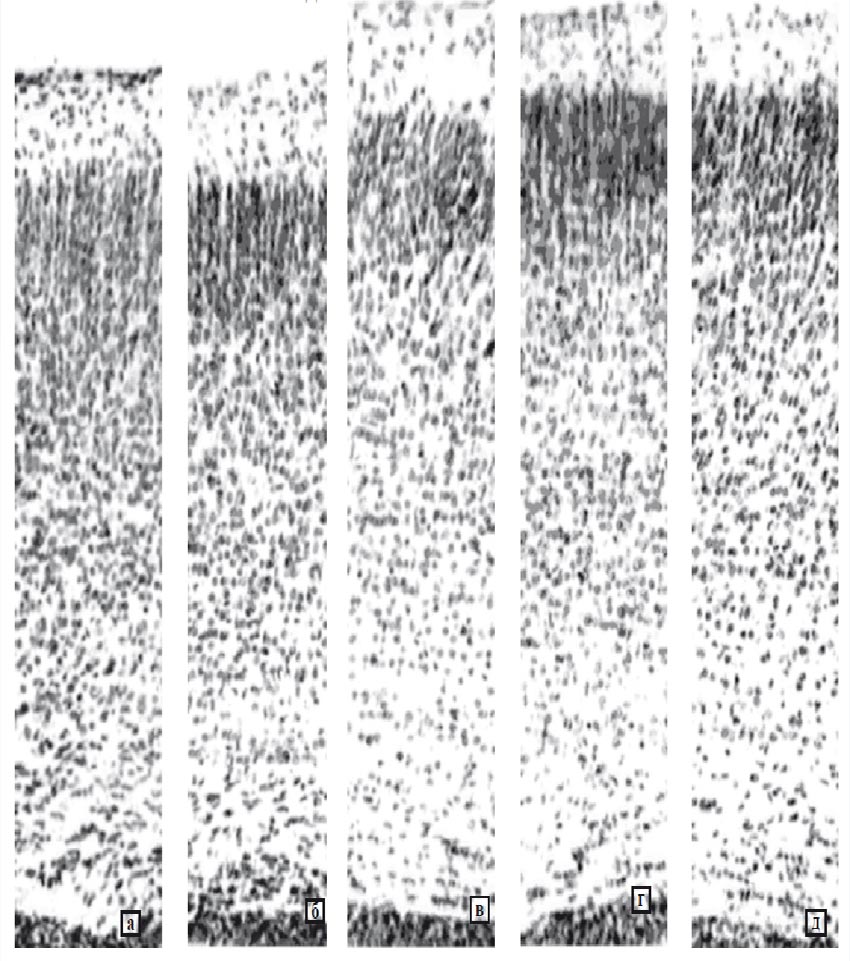
Рис. 1. Кора СТД 1-дневных крыс: потомства интактных (а), получавших масло (б), получавших прогестерон на 2 нед. беременности (25 мг/кг (в), получавших прогестерон на 3 нед. беременности (12,5 мг/кг (г); 25 мг/кг (д). Окраска галлоцианином. Увеличение 10х10
Таким образом, введение беременным крысам масла как во 2, так и в 3 нед. беременности отразилось на исследованных характеристиках мозга их однодневного потомства, причем эти различия, во-первых, были однотипными и, вовторых, более выраженными у самцов. Оценивая причины возникновения описанных отличий, можно предполагать, что они являются следствием стрессорного воздействия инъекций масла. Основанием для такого предположения служат данные литературы, свидетельствующие о том, что различные стрессорные агенты, действующие во время беременности, отражаются на пренатальном развитии различных органов, в том числе головного мозга [7, 9, 10, 12]. В связи с вышеизложенным, при изучении мозга потомства самок, получавших прогестерон, мы сопоставляли исследованные показатели как с показателями, характеризующими мозг потомства интактных животных, так и мозг потомства самок, которым вводили масло. Мозг животных группы с введением прогестерона (25 мг/кг) на 2 нед. беременности имел следующие отличия от мозга животных контрольных крысят (введение масла). У самцов наблюдалось достоверное увеличение массы органа и полушария, толщины коры ПТД и СТД, слоя I СТД. В то же время, число нейронов в стандартной площади среза слоя V было уменьшено как в ПТД, так и СТД, а также в слое II ПТД. При этом отличия от интактного контроля, имевшиеся у животных потомства самок, получавших инъекции масла, усиливались по массе мозга и полушария, толщине коры СТД и ПТД, слоя I СТД и уменьшались по такому показателю, как плотность расположения нейронов в слое V СТД и ПТД и слое II ПТД (табл. 1). У самок достоверных отличий от контроля было меньше. Они состояли в увеличении толщины коры в ПТД и уменьшении плотности расположения нейронов в слое V ПТД. В то же время, у крысят этой группы мозг отличался от мозга интактных животных большей массой, увеличенной массой полушария, толщиной коры ПТД и слоя I в ней. При этом плотность расположения нейронов в слое II и V ПТД и СТД не имела достоверных отличий от такового у потомства интактных самок (табл. 1).
Крысята обоего пола группы с введением прогестерона в дозе 25 мг/кг на 3 нед. беременности отличались от контрольных (введение масла) достоверно меньшей массой мозга и полушария. При этом гравиметрические показатели потомства подопытных самок были близки к таковым у крысят, полученных от интактных самок (табл. 2). В то же время толщина коры в СТД (только у самцов) и слоя I в ПТД и СТД (у самцов и у самок) была достоверно большей, чем у интактных крысят, и практически не отличались от таковых у потомства контрольных самок (табл. 2). Плотность расположения нейронов у самок данной группы была достоверно меньше, чем в контроле в слое V ПТД и слое II СТД. У самцов наблюдалось достоверное уменьшение плотности расположения нейронов в слое II СТД (табл. 2). Таким образом, по исследованным показателям животные данной группы занимали "промежуточное" положение между потомством интактных самок и самок, которым инъецировали масло.
Мозг новорожденных животных группы с введением прогестерона в дозе 12,5 мг/кг на 3 нед. беременности по большинству показателей не имел достоверных отличий от мозга крысят контрольной группы (введение масла) и статистически достоверно отличался от мозга потомства интактных крыс. Так, у крысят обоего пола масса мозга, полушария в подопытной группе были большие, чем у последних. Мозг самцов, кроме того, отличался большей толщиной слоя I в ПТД и СТД; у самок - коры СТД и слоя I ПТД, а также большей плотностью расположения нейронов в слое V ПТД (табл. 2).
Обзорное изучение препаратов мозга потомства самок, которым в разные сроки беременности и в различных дозах вводили прогестерон, показало, что у подопытных крысят степень стратификации неокортекса более четко выражена, чем у контрольных и интактных животных. Особенно четко это проявлялось у потомства самок, которым прогестерон вводили на 3 нед. беременности в дозе 25 мг/кг. При этом более выраженно, чем у потомства интактных и контрольных крыс, обнаруживалась граница наружных слоев с высокой плотностью расположения нейронов и внутренних, где они находились на большем расстоянии друг от друга (рисунок).
Таким образом, введение беременным крысам масла, использованного в качестве контроля, привело к ряду отличий мозга их однодневного потомства от мозга потомства интактных самок. Эти отличия, с одной стороны, могут расцениваться как отражение ускорения темпов развития мозга (увеличение массы, толщины неокортекса и слоя I ПТД и СТД). С другой стороны, при этом плотность расположения нейронов была не только не сниженной, но, напротив, более высокой, чем у потомства интактных самок. То есть уменьшения плотности расположения нейронов, характерного для опережающих темпов развития мозга, у крысят контрольной группы не наблюдалось. Как указывалось выше, есть основания предполагать, что эти изменения - следствие стресса, вызванного введением масла беременным самкам. Об этом, в частности, свидетельствуют данные о том, что эмоциональный стресс самок обусловливает увеличение толщины неокортекса, а также морфометрические изменения нейронов, а введение крысам во время беременности преднизолона (0,3 мг/кг) увеличивает плотность расположения нейронов в слое II и V ПТД и СТД [7].
Введение беременным крысам прогестерона оказало влияние на показатели морфометрии мозга их потомства, зависящее как от времени введения препарата, так и его дозы и наиболее выраженное при его инъекции на 3 нед. беременности в дозе 25 мг/кг. Так, инъекция прогестерона в дозе 25 мг/кг на 2 нед. гестации приводит к появлению признаков большей продвинутости в развитии мозга их потомства по сравнению с контролем, проявляющейся увеличением массы мозга и его полушария, толщины неокортекса в ПТД, уменьшением числа нейронов в слое V ПТД. Введение той же дозы препарата на 3 нед. беременности приводит, напротив, к уменьшению массы мозга и полушария. Вместе с тем, у подопытных крысят и этой группы имеются признаки опережающих темпов развития мозга: увеличение толщины слоя I в ПТД и (только у самцов) увеличение толщины неокортекса и слоя I в СТД, а также уменьшение числа нейронов в поле зрения в слое II СТД и (только у самок) уменьшение плотности расположения нейронов в слое V ПТД. При введении 12,5 мг/кг прогестерона на 3 нед. беременности большинство исследованных показателей не отличается от таковых у контрольных животных. В то же время, мозг подопытных крысят, по сравнению с интактными, имел признаки ускорения темпов его развития: большую массу, толщину слоя I в ПТД и СТД у самцов, увеличение толщины коры СТД и слоя I ПТД у самок. При этом не наблюдалось отличительного для опережающих темпов развития уменьшения числа нейронов в поле зрения, а, наоборот, выявлено его увеличение в слое II ПТД у самцов и слое V ПТД - у самок.
Выводы |
|
|
Из полученных нами данных следует, что введение прогестерона беременным крысам оказывает антистрессорное действие на мозг их потомства, которое более выражено у самцов, что проявляется наличием у них большего количества достоверных отличий в показателях развития мозга от таковых в контроле. Наличие эффектов, расцениваемых как антистрессорные, может рассматриваться как проявление нейропротекторного действия прогестерона.
Литература |
|
 1. Де Линьер Б. Натуральный прогестерон и его особенности // Рос. вестник акушера-гинеколога. - 2003. №6. - С. 27-30.
1. Де Линьер Б. Натуральный прогестерон и его особенности // Рос. вестник акушера-гинеколога. - 2003. №6. - С. 27-30.- 2. Ерофеев Б.Б., Иозефсон С.А., Ерофеева Л.Г. Содержание гормонов фетоплацентарного комплекса при угрожающих преждевременных родах // Дальневост. мед. журнал. - 2009. - №4. - С. 69-71.
- 3. Карева Е.Н., Олейникова О.М., Панов В.О. и др. Гестагены и головной мозг // Вестник Рос. академии мед.
- 4. наук. - 2010. - №6. - С. 40-49.
- 5. Кисели Д. Практическая микротехника и гистохимия. - М.: Медицина, 1965. - 400 с. 5. Князькин И.В., Кветной М.В., Зезюлин П.Н. и др. Нейроиммуноэндокринология мужской половой системы, плаценты и эндометрия. - СПб.: Изд-во общества "Знание" СПб. и Ленингр. обл., 2007. - 192 с.
- 6. Кобозева Н.В., Гуркин Ю.А. Перинатальная эндокринология: рук-во для врачей. - Л.: Медицина, 1986. - 311 с.
- 7. Николаева И.В. Влияние длительного стресса и введения производных гормонов коркового вещества надпочечников на показатели развития головного мозга крыс: автореф. дис. … канд. биол. наук. - Владивосток, 2009. - 23 с.
- 8. Резников А.Г. Прогестагены, беременность и здоровье плода // Гинекология. - 2003. - Т. 5, № 6. - С. 260-262.
- 9. Резников А.Г. Перинатальная модификация развития нейроэндокринной системы: феномены и механизмы // Проблемы эндокринологии. - 2004. - Т. 50, №4. - С. 42-48.
- 10. Рыжавский Б. Я. Развитие головного мозга: отдаленные последствия влияния некомфортных условий. 3-е изд., доп. - Хабаровск: Изд-во ДВГМУ, 2009. - 278 с.
- 11. Сидельникова В.М. Эндокринология беременности в норме и при патологии. - М.: МЕДпресс-информ, 2009. - 352 с.
- 12. Цывьян П.Б., Ковтун О.П. Внутриутробное программирование заболеваний детей и взрослых // Успехи физиологических наук. - 2008. - Т. 39, № 1. - С 68-75.
- 13. Brinton R.D., Thompson R.F., Foy M.R. et al. Progesterone receptors: form and function in brain // Front. Neuroendocrinol. - 2008. - Vol. 29 (2). - P. 313-339.
- 14. Schumacher M., Guennoun R., Stein D. G. et al. Progesterone: therapeutic opportunities for neuroprotection and myelin repair // Pharmacol. Ther. - 2007. - Vol. 116. - P. 77-106.
 |
Главное меню |
|
Заглавие |
|
Введение |
|
Материалы и методы |
|
Результаты и обсуждение |
|
Выводы |
|
Литература |
Оригинальная верстка  |
|